
Ali M. Izzo 1*; Hassan Khojah 2; Abdul-Mohsen Murie 1
1, General Commission for Scientific Agricultural Research (GCSAR), Syria
2, Department of Horticulture, Faculty of Agriculture, Tishreen University, Latakia, Syria
E-mail:
izzo198899@gmail.com
Received: 17/07/2021
Acceptance: 28/10/2021
Available Online: 01/11/2021
Published: 01/01/2022

Manuscript link
http://dx.doi.org/10.30493/DAS.2021.295501
Abstract
This study was conducted to estimate heterosis and combining ability for yield and some economic-related characteristics in tomato. For this purpose, six tomato lines with their 15 direct hybrids resulted from half-diallel mating were cultivated during the 2017 and 2018 seasons in Al-Jamasah station, Tartous agricultural research center, Syria. Results showed that most of the hybrids were characterized by significant desirable heterosis compared to mid-parents and the best-parent for most of the studied traits. Heterosis reached 49.03 and 33.4% for single plant yield, -31.2 and -29.27 for the number of days to the beginning of flowering, 89.73 and 61.77% for the number of fruits per cluster, 15.72 and 9.30% for fruit height, 27.29 and 24.18% for fruit diameter, 32.55 and 24.18% for the number of locales per fruit comparing to mid-parents and best-parent, respectively. The hybrids T2×T6, T2×T8, and T8×T16 were significantly superior and could be promising hybrids for improving yield potential. Variations related to general and specific combining ability were highly significant, indicating the role of additive and non-additive gene action in the inheritance of the studied traits. Furthermore, the ratio (σ2GCA/σ2SCA) showed the control of additive gene action in the inheritance of fruit weight, fruit height, fruit diameter, and the number of fruits per cluster, while non-additive gene action controlled the inheritance of single plant yield, the number of days to flowering and the number of locales per fruit.
Keywords: Solanum Lycopersicum, General combining ability, Specific combining ability, Diallel, Hybrid
Introduction
Tomato is one of the most important vegetable crops widespread all over the world. The global area planted with tomatoes reached 5.03 million hectares, and the total production reached 180.8 million tonnes [1]. In Syria, tomato is produced under protected as well as non-protected conditions. In 2018, the total cultivated area of this crop was 3878 ha, and the total production was 581754 tons [2]. Tomato is characterized by high nutritional value and multiple industrial usages [3].
Genetic improvement of tomato aimed to produce hybrids characterized by high yield potential and enhanced fruit quality. Heterosis evaluation is a crucial step in genetic improvement programs, which depends on choosing the suitable parents for hybridization to guarantee the production of promising hybrids characterized by superior traits [4]. Several authors mentioned the importance of information related to the type of gene action that controlled the economic traits and combining ability in the success of genetic improvement of tomato. This information helps breeders to select suitable parents and consequently the most effective breeding strategy [5].
Previously, high significant variation in general and specific combining ability was reported in tomatoes [6]. This finding indicated the role of additive and non-additive gene action in the inheritance of the studied traits. Additionally, the ratio σ2GCA/σ2SCA highlighted theadditive gene control action in the inheritance of fruit weight, the number of locales per fruit, and plant yield. Combining ability and gene action of yield and other related traits such as fruit weight, number of days until the start of flowering, number of locules per fruit, number of fruit per plant, total sugars, titrable acidity, and total soluble solids was previously studied [7]. The results showed highly significant variation in general and specific combining ability for most studied traits, which indicated the role of additive and non-additive gene action in the inheritance of the studied traits. Furthermore, the σ2GCA/σ2SCA ratio referred to thenon-additive gene action in the inheritance of all traits. Relatively low to medium mid-parents and best-parent heterosis were reported for yield and other related traits [8], while slightly higher heterosis results were observed for the same traits in [9].
The present study aimed to analyze the genetic behavior of yield and fruit traits using heterosis, general and specific combining ability of some local tomato inbred lines. The obtained results could determine the superior hybrids with commercial potential or be used in future tomato breeding programs.
Materials and Methods
Study site
The study was conducted in Tartous center for scientific agricultural research – Al-Jamaseh station.
Plant material
Six lines of tomato bred in the general commission for scientific agricultural research have been used in this study. Those lines are characterized by high variation in growth habits, earliness, and fruit traits (Table 1). Seeds were planted in trays and then transplanted after 21 days. Half-Diallel (excluding reciprocal) hybridization between the lines was carried out in the season of 2017. The seeds of the 15 hybrids and the parental lines were planted in the greenhouse in 2018 season in a completely randomized block design (RCBD) with three replications. The planting distance was 90 cm between rows and 40 cm between the plants in the same row. All agricultural practices were done according to GCSAR recommendations for greenhouse tomatoes.
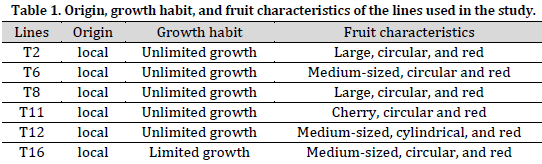
Studied traits
Single plant yield, days from transplanting to the start of flowering, average fruit weight, number of fruits per cluster, number of locules per fruit, fruit diameter, and fruit height were taken from eight surrounded plants in each experimental plot.
Data analysis
GenStat 10 was used for statistical analysis. The significance of heterosis compared to the mid and best parent was estimated using the t-student test [10]. General combining ability (GCA) and specific combining ability (SCA) were estimated using the second method and the first model of Griffing [11].
Results and Discussion
Variations in lines and hybrids
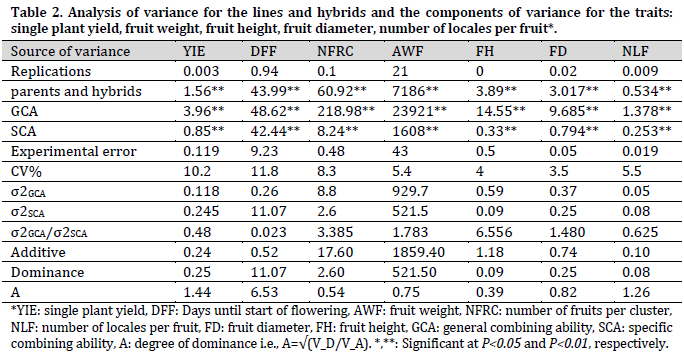
The parental lines had a highly significant variation for all the studied traits, indicating genetic distance among the parental lines for the studied traits (Table 2).
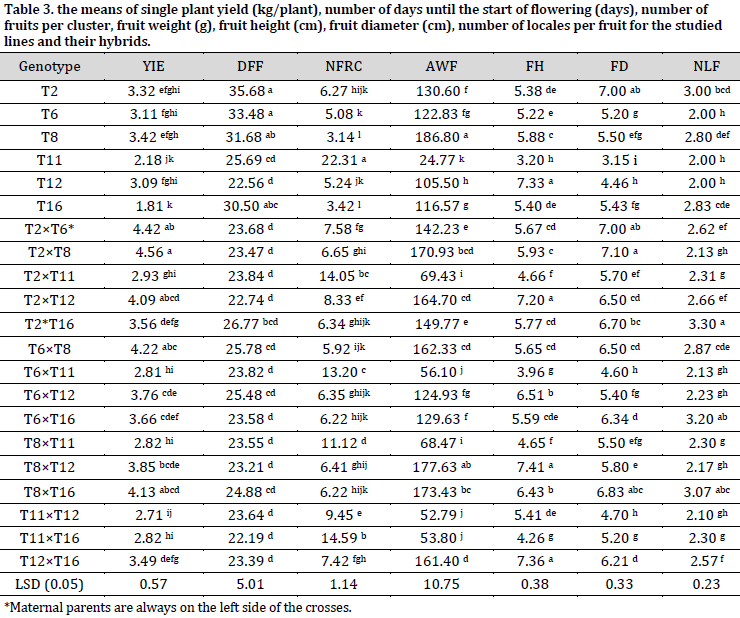
Regarding single plant yield (kg/plant), T8 had the highest single plant yield (3.42 kg/plant), while T16 had the lowest (1.81 kg/plant). A significant variation in the number of days until the start of flowering among the studied lines was found, with T12 being the earliest line (22.56 days), while T2 was the latest (35.68 days). The studied lines showed significant differences in the number of fruits per cluster, ranging from 3.14 to 22.31 fruit per cluster in T8 and T11, respectively.
Average fruit weight ranged between 186.80 g (T8) and 24.77 g (T11). Significant differences were found among the lines in terms of fruit height and diameter. The highest fruit height was observed in line T12 (7.33 cm), and T2 recorded the highest fruit diameter (7.00 cm), while line T11 scored the lowest fruit height and diameter with 3.20 cm and 3.15 cm, respectively. Significant differences in the number of locules per fruit were also found, ranging between 2 and 3 locales per fruit (Table 3).
The hybrids had significant variation in their characteristics, confirming the genetic distance of the parental lines used in hybridization. For single plant yield, the means of the hybrids varied between 2.71-4.56 kg/plant. The highest yield was recorded in the hybrid (T2×T8), and the lowest was for (T11×T12). The hybrids differed in the number of days until flowering; the hybrid (T11×T16) was the earliest (22.19 days), while the (T2×T16) was the latest (26.77 days). Significant differences in the number of fruits per cluster were found among the hybrids. (T11×T16) had the highest number of fruits per cluster (14.59 fruit/cluster), while (T6×T8) had the lowest (5.92 fruit/cluster). As for average fruit weight (g), (T2×T8) gave the fruit with the highest weight (170.93 g), while (T11×T12) gave the lowest (52.79 g).
The hybrids showed significant differences in fruit height (cm), which ranged between 7.41 cm (T8×T12) and 3.96 cm (T6×T11). For fruit diameter (cm), the hybrid (T2×T8) gave the highest fruit diameter (7.10 cm), while the hybrid (T6×T11) gave the lowest (4.60 cm). The number of locales per fruit ranged between 2.10 locales per fruit for the hybrid (T11×T12) and 3.30 for the hybrid (T2×T6) (Table 2). These results are similar to [12], where significant variations among 30 genotypes in the number of flowers per cluster, number of fruits per cluster, fruit weight, and single plant yield were reported. Additionally, [5] showed significant variation among half-diallel hybrids and their parents in terms of yield, fruit weight, and the number of locules per fruit.
Heterosis
Most hybrids showed significant positive heterosis for single plant yield (Table 4). Heterosis values ranged between 18.28-49.03% and between 21.01-33.40% compared to the mid-parents and the best-parent, respectively.
In terms of the number of fruits per cluster, eleven hybrids showed significant positive desirable heterosis compared to the mid-parents ranging between 13.41-89.73%. Also, six hybrids showed significant positive desirable heterosis compared to the best parent, and the highest heterobeltiosis was recorded in the hybrid (T8×T16) (81.77%) (Table 4).
For the number of days until flowering, eleven hybrids showed desirable significant negative heterosis ranging between -17.91 and -31.52% compared to the mid-parents, while five hybrids showed desirable significant negative heterobeltiosis. The most negative values for both heterosis indicators were noted in the hybrid (T2×T6) with -31.52 and -29.27 compared to the mid-parents and best parent, respectively (Table 4).
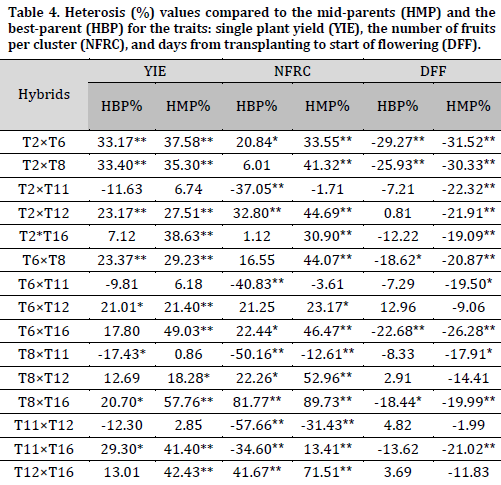
As for average fruit weight, nine hybrids showed significant positive desirable heterosis compared to the mid-parents, ranging between 8.30-45.36%. The highest heterosis was recorded in the hybrid (T12×T16). Four hybrids showed significant positive desirable heterosis compared to the best parent. The highest heterobeltiosis value was for the hybrid (T12×T16) (38.46%) followed by (T2×T12) (26.11%) (Table 4).
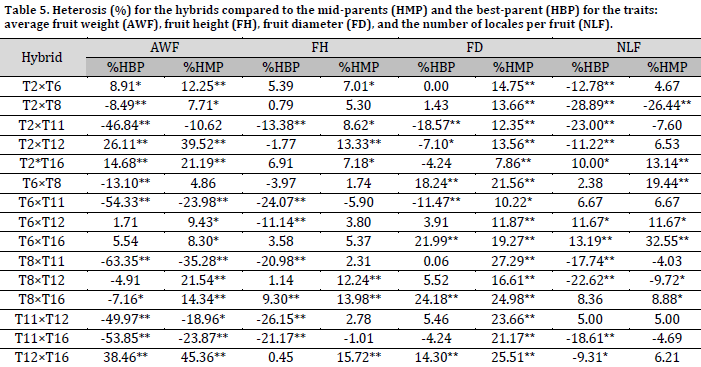
Regarding fruit height, seven hybrids showed significant positive heterosis compared to the mid-parents. The highest heterobeltiosis was for the hybrid (T12×T16) (15.72%) (Table 5). All studied hybrids possessed significant positive heterosis in terms of fruit diameter compared to the mid-parents. The highest heterosis value was for the hybrid (T8×T11) (27.29%).
Five hybrids showed significant positive desirable heterosis compared to the mid-parents for the number of locules per fruit. The highest heterosis was for the hybrid (T6×T16) (32.55%). Three hybrids showed significant positive heterosis compared to the best parent with the highest value (13.19%) recorded in the hybrid (T6×T16) (Table 5). Similar results were obtained by [9], who reported significant heterosis reaching 62.5% for single plant yield, -25% for the number of days until the flowering of 50% of the flowers, and 41.2% for fruit weight. In another study, heterosis values of 14.2, 12.26% for the number of fruits per cluster, 13.75, 11.47 for fruit height, 21.49, 10.24 for fruit diameter, 29.65, 26.7 for fruit weight, and 34.73, 31.82 for single plant yield compared to mid-parents and best-parent, respectively [13]. Similarly, [14] reported significant heterosis for the number of days to flowering (-14.64%) and fruit weight (15.8%) compared to the best parent.
Combining ability
Significant general and specific combining ability values were observed in all studied traits (Table 2). This indicates the role of additive and non-additive gene action in the inheritance of those traits. A σ2GCA/σ2SCA ratio of 0.48 was recorded in single plant yield, which indicates the importance of non-additive gene action in the inheritance of this trait. Furthermore, the degree of dominance (a=1.44) confirmed this gene behavior. These results are in accordance with [15-21]. The ratio (σ2GCA/σ2SCA=0.023) and the degree of dominance (a=6.53) showed the important role of non-additive gene action in the inheritance of days to flowering. This finding was in accordance with the results of [7][22].
In the number of fruits per cluster, additive gene action had the highest role in the inheritance of this trait since high σ2GCA/σ2SCA ratio (3.385) and low dominance (0.54) values were observed, which meets the results of [22][23]. Similarly, fruit weight, height, and diameter seem to be controlled by additive gene action since these traits scored σ2GCA/σ2SCA ratios of 1.783, 6.556, and 1.480 and dominance values of 0.75, 0.39, and 0.82, respectively. These results are in accordance with [23-25]. This genetic control of additive nature can be utilized in improving those traits by selection in the later filial generations [17].
Conversely, non-additive gene action controlled the inheritance of the number of locales per fruit since a low σ2GCA/σ2SCA ratio (0.625) and high dominance (1.26) were observed. This was in accordance with [6][7][14][16][17][24]. Therefore, the non-additive gene action controlled the inheritance of single plant yield, number of days until the start of flowering, and number of locales per fruit. Thus, heterosis could be desirable in improving those traits [15][20].
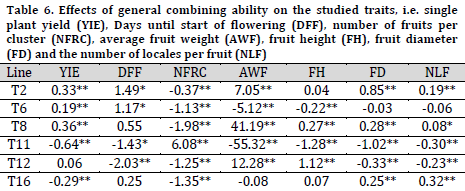
Single plant yield GCA effects in T8 (0.36), T6 (0.19), T2 (0.33) were significantly positive. Therefore, those lines could be used as a donor of this trait (Table 6). Regarding the number of days until the start of flowering, which is an index of earliness, T11 and T12 scored the highest significant negative effects (-1.43 and -2.03). Thus, these two lines could be used in producing early maturing hybrids [7][22]. As for the number of fruits per cluster, T11 scored a significant positive GCA, while other lines had significant negative GCAs. Based on these results, T11 could be helpful in improving the number of fruits per cluster. T8 scored the highest significant positive GCA effects for average fruit weight, followed by T12 and T2. In fact, T8 can be suggested in breeding programs to introduce fruits with higher weights, heights, and diameters (higher volume), while T2 is suggested to produce types with higher fruit weight and wider diameter [5][22][25].
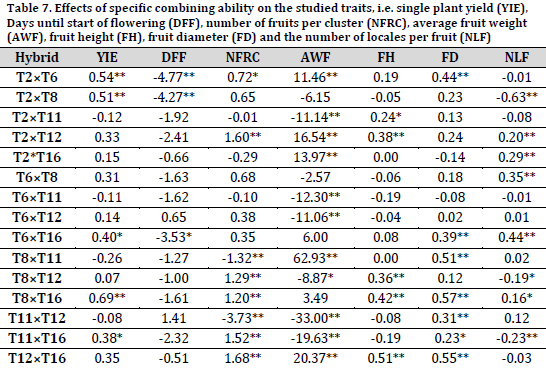
Five hybrids showed significant positive desirable specific combining ability on single plant yield, mainly the hybrids (Table 7). T2×T6 and T2×T8 recorded the most negative SCA values in term of earliness of flowering. For the number of fruits per cluster, six hybrids showed significant positive desirable combining ability. T8×T16 scored significant positive SCAs for fruit weight, hight, and diameter. Those hybrids could be used in improving the studied traits [26].
Conclusions
The studied lines differed in their characteristics and general combining ability; many of them could be used to improve one or more of the studied traits. High heterosis was achieved in most of the studied traits. Additive gene action controlled the inheritance of average fruit weight, fruit height, fruit diameter, and the number of fruits per cluster. Therefore, it is recommended to select in the mid and later generations for improving those traits. On the other hand, non-additive gene action controlled the inheritance of single plant yield, number of days until flowering, and the number of locales per fruit; thus, selection could be made in early generations to improve those traits. T2 and T8 could be introduced in breeding programs since they possess a desirable general combining ability for most studied traits. Additionally, the hybrids T2×T6, T2×T8, and T8×T16 are promising in improving fruit yield for their high SCA and heterosis in this trait.
References
| 1 | FAO. Plant genetic resource for food and agriculture. Rome, Food and Agriculture Organization of the United Nations.2019. |
| 2 | Syrian Ministry of Agriculture and Agrarian Reform (MOAAR). Directorate of Planning and Statistics, Division of Statistic. Syria. 2018. |
| 3 | Maurya RK, Singh AK, Sai A. Correlation and path analysis in tomato (Solanum lycopersicum L.) for yield and yield contributing traits. J. pharmacogn. phytochem. 2020;9(3):1684-7. |
| 4 | Hannan MM, Biswas MK, Ahmed MB, Hossain M, Islam R. Combining ability analysis of yield and yield components in tomato (Lycopersicum esculentum Mill.). Turk. J. Botany. 2007;31(6):559-63. |
| 5 | Muttappanavar RD, Sadashiva AT, Vijendrakumar RC, Roopa BN, Vasantha PT. Combining ability analysis of growth, yield and quality traits in cherry tomato (Solanum lycopersicum var. cersiforme). Mol. Plant Breed. 2014;5. |
| 6 | Farzane A, Hossein N, Hossein A, Mirshamsi KA, Navid V. The Estimate of Combining Ability and Heterosis for Yield and Yield Components in Tomato (Lycopersicon esculentum Mill.). J. Biol. Environ. Sci. 2012;6(17):129-34. |
| 7 | Dharva PB, Patel AI, Vashi JM, Chaudhari BN. Combining ability analysis for yield and yield attributing traits in tomato (Solanum lycopersicum L.). Int. J. Chem. Stud. 2018; 6(3):2342-8. |
| 8 | Purkayastha MD, Mahanta CL. Physicochemical properties of five different tomato cultivars of Meghalaya and their suitability in food processing. Afr. J. Food Sci.,2011; 5(12):657-67. |
| 9 | Soresa DN, Nayagam G, Netsanet B, Jaleta Z. Heterosis in Tomato (Solanum lycopersicum L.) for Yield and Yield Component Traits. Adv. Res. 2020;21(9):141-152. DOI |
| 10 | Sinha SK, Khanna R. Physiological, biochemical, and genetic basis of heterosis. InAdvances in Agronomy. Academic Press. 1975;27:123-74. DOI |
| 11 | Griffing B. A generalised treatment of the use of diallel crosses in quantitative inheritance. Heredity. 1956;10(1):31-50. DOI |
| 12 | Najeema NH, Hadimani RHP, Biradar IB. Evaluation of Cherry Tomato (Solanum lycopersicum var. cerasiforme) Genotypes for Yield and Quality Traits. Int. J. Curr. Microbiol. App. Sci. 2018; 7(6):2433-9. DOI |
| 13 | Kumar P, Paliwa A, Tiwari D, Upadhyay S, Bahuguna P. Heterosis Studies For Yield Andits Attributing Traits In Tomato Under Mid-Hill Conditions Of Garhwal. Int. J. Adv. Res. 2016;4(11):750-7. DOI |
| 14 | Gautam N, Kumar M, Kumar D, Kumar S, Vikram A, Dogra RK, Sharma S. Combining ability and gene action studies for important quality traits in tomato (Solanum lycopersicum L.). Int. J. Chem. Stud. 2018;6(2):1992-6. |
| 15 | Kansouh AM, Zakhe AG. Gene Action And Combining Ability In Tomato (Lycopersicon Esculentum Mill.) By Line X Tester Analysis. J. Plant Production .2011;2(2):213–27. DOI |
| 16 | Solieman THI, El-Gabry MAH, Abido AI. Heterosis, potence ratio and correlation of some important characters in tomato (Solanum lycopersicum L.). Sci. Hortic. 2013;150:25-30. DOI |
| 17 | Shankar A, Reddy R, Sujatha M, Pratap M. Combining ability analysis to identify superior F1 hybrids for yield and quality improvement in Tomato (Solanum lycopersicum L.). Agrotechnology. 2013;2(114). DOI |
| 18 | Agarwal A, Arya DN, Ranjan R, Zakwan A. Heterosis, combining ability and gene action for yield and quality traits in tomato (Solanum lycopersicum L.). Helix. 2014;2:511-5. |
| 19 | El-Gabry MAH, Solieman TIH, Abido AIA. Combining ability and heritability of some tomato (Solanum lycopersicum L.) cultivars. Sci. Hortic. 2014;167:153–7. DOI |
| 20 | Gayosso-Barragán O, López-Benítez A, Rodríguez-Herrera SA, Ek-Maas JN, Hidalgo-Ramos DM, Alcala-Rico JS. Studies on combining ability in tomato (Solanum lycopersicum L.). Agron. Res. 2019; 17(1):77–85. DOI |
| 21 | Vekariya T, Kulkarni GU, Vekaria DM, Dedaniya AP, Memon JT. Combining ability analysis for yield and its components in tomato (Solanum lycopersicum L.). Acta sci. agric. 2019;3(7):185-91. DOI |
| 22 | Mishra A, Nandi A, Sahu GS, DAS S, Mohanty IC, Pattanayak SK, Tripathy P. Studies of combining ability in tomato (Solanum lycopersicum L.) for vegetative growth, yield and quality traits. J Pharmacogn Phytochem. 2020;9(1):466-73. |
| 23 | Ahmad S, Quamruzzaman AKM, Nazim Uddin M. 2009. Combining Ability Estimates of Tomato (Solanum Lycopersicum) In Late Summer. SAARC J. Agri.2009;7(1),43-56. |
| 24 | Garg N, Cheema DS, Dhatt AS. Genetics Of Yield, Quality And Shelf Life Characteristics In Tomato under normal and late planting conditions. Euphytica. 2008;159:275–88. DOI |
| 25 | Aisyah SI, Wahyuni S, Syukur M, Witono JR. The Estimation of Combining Ability and Heterosis Effect for Yield and Yield Components in Tomato (Solanum lycopersicum Mill.) at Lowland. Ekin J. Crop Breeding and Genetics.2016; 2(1):23-29. |
| 26 | Raj T, Bhardwaj ML, Pal S, Kumrai S, Dogra RK. Performance of Tomato (Solanum lycopersicum L.) Hybrids for Yield and its Contributing Traits under Mid-hill Conditions of Himachal Pradesh. Int. J. Bio-resour. Stress Manag. 2018;9(2):282-6. DOI |
Cite this article:
Izzo, A., Khojah, H., Murie, A. Combining ability and heterosis for yield and some fruit traits of tomato. DYSONA – Applied Science, 2021;3(1): 15-23. doi: 10.30493/das.2021.295501